| |
Plasma membrane
& transport, Organelles involved in metabolism
Filtration: Imagine
water (interstitial fluid) dripping through a coffee filter (Semi-permeable
membrane) and depositing only small dissolved (diffusable) molecules
into your cup (cell). Try this:
Blood pressure (water / hydrostatic pressure) leaking through capillary
cells (epithelial cells) and leaving nutrients (glucose, etc) for
your cells.
Simple Diffusion:
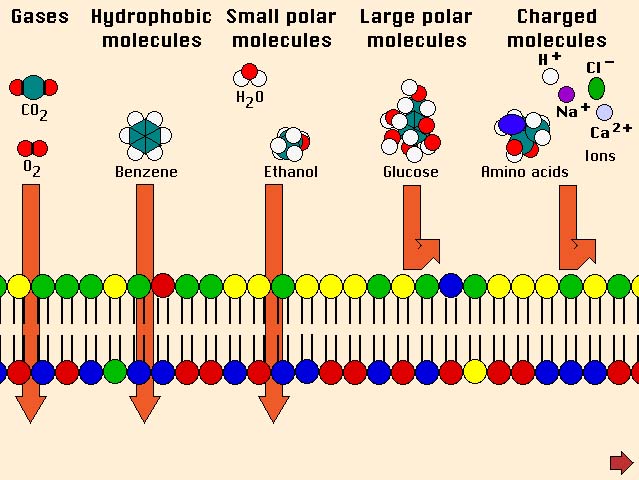
Water, oxygen, carbon dioxide, ethanol and urea are examples of
molecules that readily cross cell membranes by simple diffusion.
They pass either directly through the lipid bilayer or through pores
created by certain integral membrane proteins. The relative rate
of diffusion is roughly proportional to the concentration gradient
across the membrane. For example, oxygen concentrations are always
higher outside than inside the cell and oxygen therefore diffuses
down its concentration gradient into the cell; the opposite is true
for carbon dioxide.
The limiting step for simple diffusion across a cell membrane is
movement of the molecule from the aqueous environment outside or
inside the cell into the lipid bilayer
of the membrane. Rate of transport for a particular
molecule is therefore proportional to the lipid solubility or hydrophobicity
of that molecule. Oxygen, carbon dioxide
and ethanol are highly lipid soluble and therefore diffuse across
the bilayer almost as if it were not there.
Water, however, is
not at all lipid soluble (almost by definition) and is polar.
However, it seems that water is small enough and has sufficient
kinetic energy that it can diffuse through the lipid bilayer with
minimal interference. Certain other small, uncharged, hydrophilic
molecules, urea being an example, diffuse across lipid bilayers
with relative ease.
Ions and charged molecules diffuse
cross the lipid bilayer of cell membranes very, very poorly.
Their charge, either positive or negative, causes them to be repelled
from like charges in the cell membrane. Additionally, their charge
causes them to electrically bind water molecules, causing them to
be hydrated and effectively quite large.
To summarize,
many small and uncharged molecules diffuse across the plasma membrane
by virtue of their kinetic energy
of motion. In most
cases, such molecules are hydrophobic, nonpolar and can dissolve
in and out of membrane lipid. Bearing a charge or having a large
size virtually abolishes an ability to be transported across lipid
membranes by simple diffusion, but do not fear - membranes have
other means to facilitate
such transport.
Osmosis: If
two solutions of different concentration are separated by a semi-permeable
membrane which is permeable to to the smaller solvent molecules
but not to the larger solute molecules, then the solvent will tend
to diffuse across the membrane from the less concentrated to the
more concentrated solution. This process is called osmosis.
Osmosis is of great importance in biological processes where the
solvent is water. The transport of water and other molecules across
biological membranes is essential to many processes in living organisms.
The energy which drives the process is usually discussed in terms
of osmotic pressure.

| |
| Click: Osmolarity
and Tonicity |
|
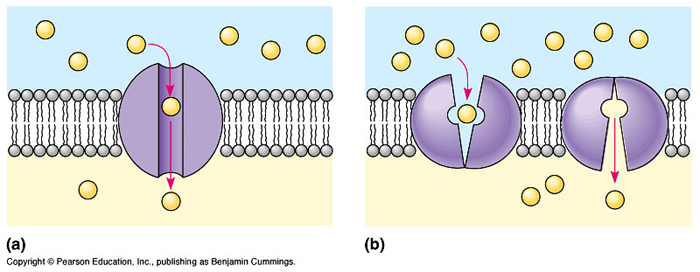
Carrier-Mediated
Transport
Facilitated "diffusion"
- The use of the term "diffusion"
in conjunction with this carrier-mediated process is unfortunate
because it often implies that the substrate crosses the membrane
owing only to the passive permeability characteristics of
the membrane. In fact, the use of the term merely serves to
emphasize that this process is only capable of a net movement
of solute across a membrane in response to a trans-membrane
electrochemical gradient ( high
to low concentration ) of the substrate molecule.
For example, glucose crosses the membrane of virtually every
cell in the body by facilitated diffusion. The process results
in the movement of glucose from high concentrations in the
plasma to lower concentration in the cell; it cannot produce
a higher concentration in the cell than that of the surrounding
solution, and in that respect it is similar to simple diffusion.
However, the family of membrane proteins responsible for catalyzing
the process (the GLUT family of transport proteins) obeys
Michaelis-Menten type kinetics, shows structural specificity,
and can be inhibited by specific toxins. Cellular metabolism
can result in facilitated diffusion by continuously using
up the substrate after it enters a cell, thereby maintaining
the concentration gradient.
Active transport
uses energy (in the form of ATP), and materials flow against
the concentration gradient. Carrier-mediated
active transport systems use energy and membrane
proteins to "pump" certain substances against a
concentration gradient. This causes the substance to accumulate
on one side of the plasma membrane. An example of this type
of active transport is the sodium-potassium pump, which consists
of a membrane protein that uses ATP to pump Na+ (sodium) out
of the cell and K+ (potassium) into the cell. These unbalanced
accumulations of Na+ and K+ are needed for proper nerve functioning.
|
Vesicular Transport
Intracellular vesicle traffic is crucial
for normal cell function in all eukaryotic cells, from yeast
to human. Transport of proteins within the extensive network
of membrane-bound compartments is highly regulated to ensure
the specificity and efficiency of cargo delivery.
Vesicular transport does not require the protein molecules
to pass through membranes. Instead it is the membrane that
migrates and fuses with other compartments taking the protein
along with it, via a process known as pinocytosis.
Movement between the endoplasmic
reticulum and the Golgi
apparatus occurs in this manner.
Example 'A' shows receptors
for LDL (which binds cholesterol).
Recall that LDL is considered the "bad"
cholesterol because its role is to bring cholesterol
into the cell so it can be used. If it is high in the blood
stream, this means that cholesterol is not being taken up,
or is too high. Sometimes this can be due to a defect in the
LDL receptor. In A, above, note that the LDL receptor + cholesterol
move to clathrin coated pits thanks to Adaptin which binds
the receptor and the clathrin. Then, they move from early
to late endosomes. Normally, vesicles
then take the cholesterol to the Golgi complex or
it is released for use by the cell.
In 'B', however
the LDL receptor has lost its adaptin binding site. This is
a genetic mutation and causes hypercholesterolinemia. The
LDL receptor can not be sequestered in the clathrin coated
pit and can not be taken up.

|
|
|